A model of a sodium channel¶
The second ion channel in Hodgkin-Huxley model controls sodium. The theory here is the basis for the the practical modelling tutorial in Tutorial 7.
Theory¶
The Hodgkin-Huxley model’s sodium channel has two types of gate, an \(m\)-gate (of which there are 3) that is initially closed (\(m = 0\)) before activating and inactivating back to the closed state, and an \(h\)-gate that is initially open (\(h = 1\)) before activating and inactivating back to the open state. The short period when both types of gate are open allows a brief window current to pass through the channel. Therefore,
where \(\bar{g}_{Na} = \ \)120 mS.cm-2, and with \(\left\lbrack Na^{+} \right\rbrack_{i}\)= 30mM and \(\left\lbrack Na^{+} \right\rbrack_{o}\)= 140mM, the Nernst potential for the sodium channel (\(z=1\)) is:
TODO … this doesn’t equal 35 …??
The gating kinetics are described by:
where the voltage dependence of these four rate constants is determined experimentally 1 to be:
Interpretation as a CellML model¶
Before we construct a model of the sodium channel, we first
introduce some further CellML concepts that help deal with the
complexity inherent in biological modelling. First we’ll look at the use of
encapsulation groups and public and private interfaces to control the
visibility of information in modular CellML components. To understand
encapsulation, it is useful to use terms like parent, child and sibling,
to describe the relationships between components. This is illustrated in
encapsulation_parent_structure below:
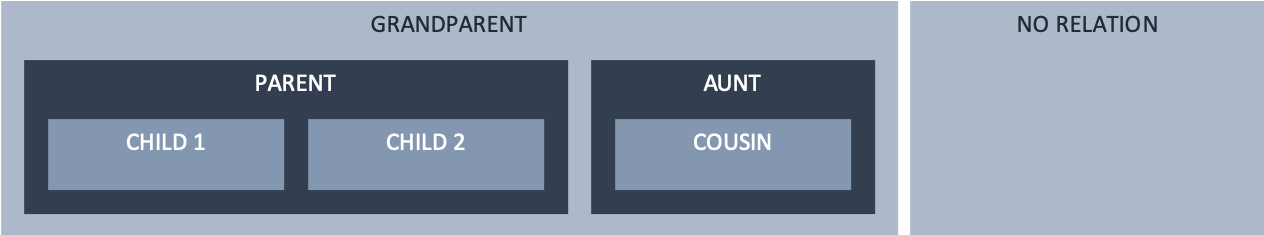
Encapsulation structure of nested components in a parent-child-sibling structure.¶
Defining an encapsulation structure controls which components are able to see and be seen by which others, as defined by the available interface types:
By default, no interface exists between components. The types of interface listed below are those which are possible; but each must be explicitly specified before it can be used. While
noneis the implied default, it may be specified explicitly as well.A
publicinterface is available between siblings. This is also true of top-level components (which are siblings with aModelparent), such as the ‘grandparent’ and ‘no relation’ components inencapsulation_parent_structure. Apublicinterface is also available from a child to its parent (note direction).A
privateinterface is available from a parent to its child(ren), again noting the direction.Both
publicandprivateinterfaces are available using thepublic_and_privatetype, needed where there is more than one level of nesting (for example, a grandparent-parent-child structure). Here, the parent must define apublic_and_privateinterface type as it is both the child of the grandparent, and the parent of the child.Communication through the aunt-child, grandparent-child, and cousin-parent relationships is not possible as there is no available interface. Communication with the no-relation component is only possible with the grandparent because, as top-level components, they are siblings.
We define the CellML components sodium_channel_m_gate and sodium_channel_h_gate below. Each of these components has its own equations (voltage-dependent gates and first-order gate kinetics) but they are both parts of one protein – the sodium channel – so it is useful to group them into one sodium_channel component.
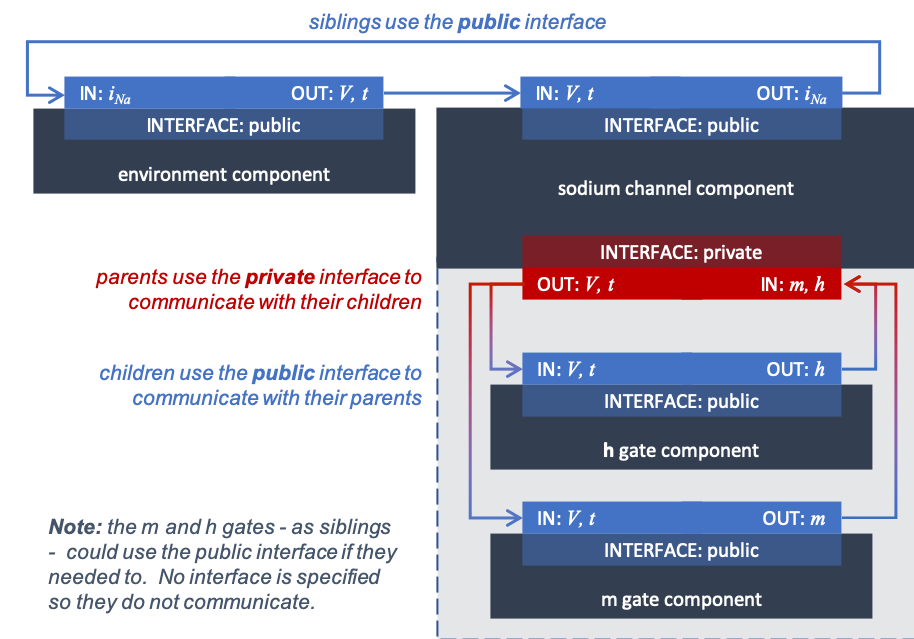
The sodium channel component is the parent of two children: the \(m\)-gate and the \(h\)-gate components, which are therefore siblings. A private interface allows a parent to talk to its children and a public interface allows siblings to talk among themselves and to their parents.¶
Simulation and results¶
The behaviour of the sodium channel was simulated using the
simple solver provided to run the code generated
in Tutorial 7. Three voltage-step experiments
were run with voltage conditions of
(a) -85mV to -20mV, (b) -85mV to 0mV and (c) -85mV to +20mV.
The results of the computation, with an end time of 40ms, and Euler step of
0.01ms are shown in sodium_v to sodium_i below.
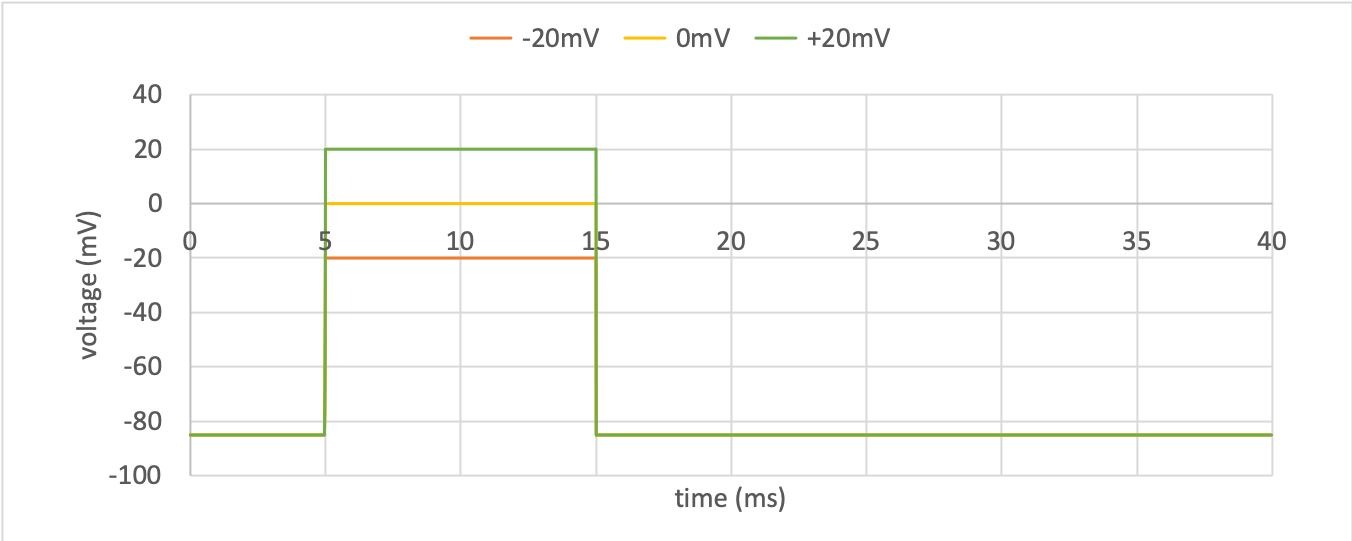
Driving function for the voltage clamps¶
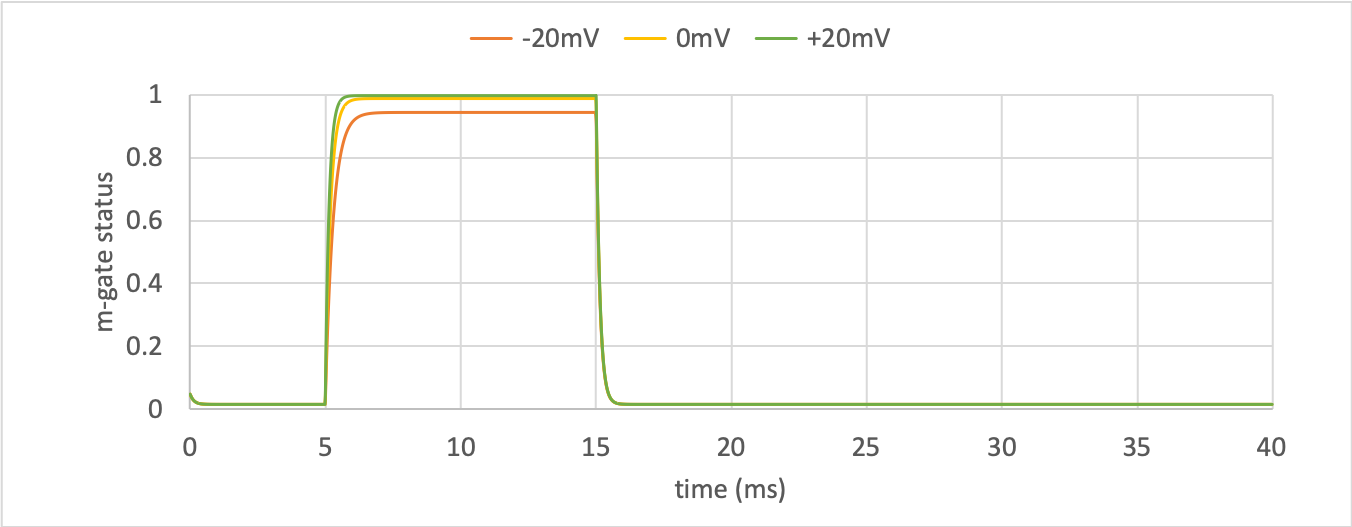
m-gate dynamics¶
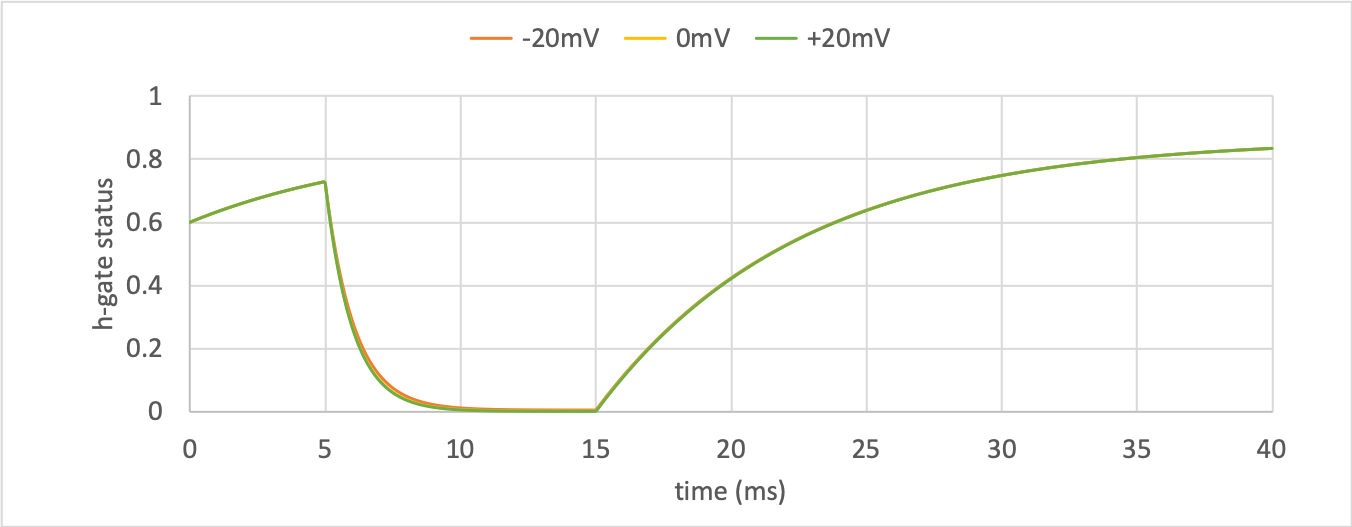
h-gate dynamics¶
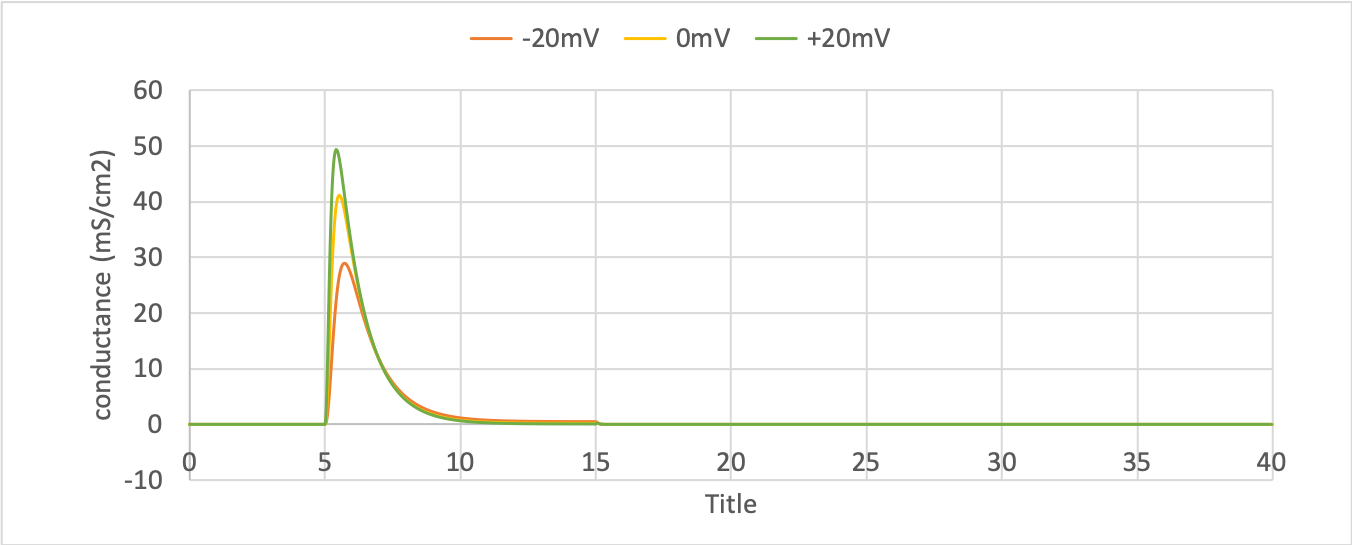
Sodium conductance¶
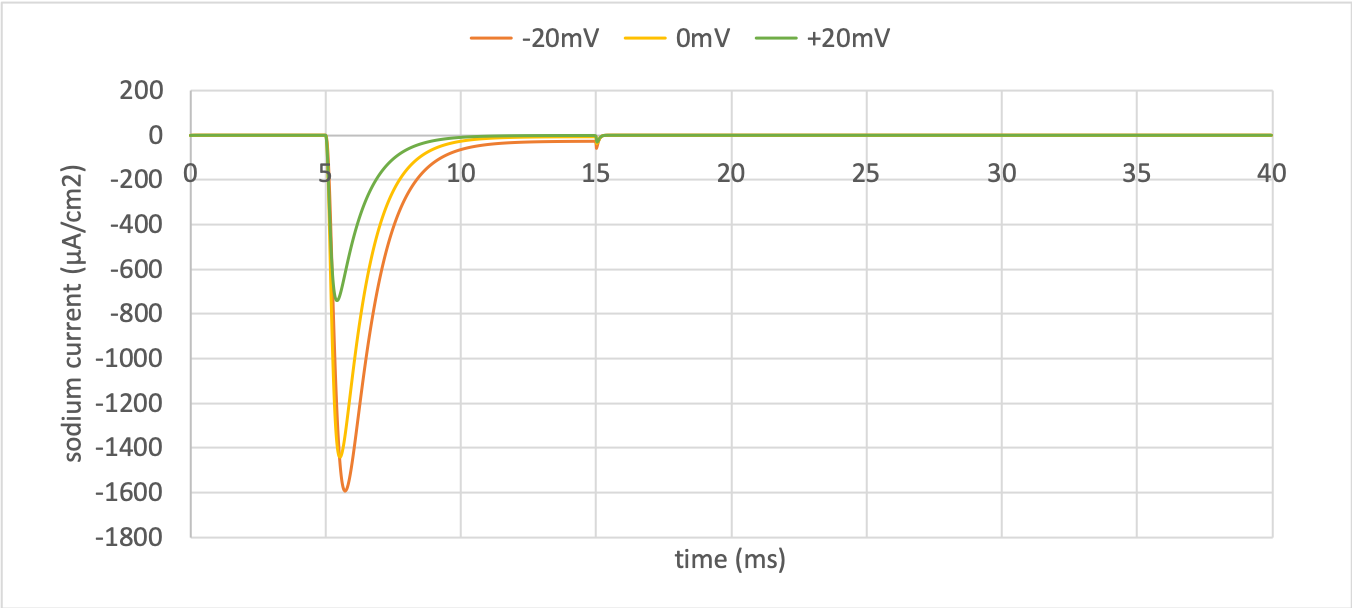
Sodium current¶
Notable results¶
Of particular note in the results are these features:
The kinetics of the \(m\)-gate are much faster than the \(h\)-gate.
The opening behaviour is faster as the voltage is stepped to higher values since \(\tau = \frac{1}{\alpha_{n} + \beta_{n}}\) reduces with increasing \(V\) (see
ocr_tut_volt_deps_gates).The sodium channel conductance rises (activates) and then falls (deactivates) under a positive voltage step from rest since the three \(m\)-gates turn on, but the \(h\)-gate turns off and the conductance is a product of these. Compare this with the kinetics of the potassium channel conductance which is only reduced back to zero by stepping the voltage back to its resting value – that is, deactivating it.
The only time current \(i_{Na}\) flows through the sodium channel is during the brief period when the \(m\)-gate is rapidly opening and the much slower \(h\)-gate is beginning to close. A small current flows during the reverse voltage step but this is at a time when the \(h\)-gate is now firmly off so the magnitude is very small.
The large sodium current \(i_{Na}\) is an inward current and hence negative.
Next steps¶
The incorporation of this sodium channel model into the full Hodgkin-Huxley model of a neuron is described in the next chapter, Hodgkin-Huxley assembly. This will include the \(n\)-gate described in the previous chapter, A model of a potassium channel as well as a leakage current model.
Footnotes
- 1
The original version of the Hodgkin-Huxley paper used: \(\alpha_m = \frac{0.1(V+25)} {\exp\left(0.1(V+25)\right)-1}\), \(\beta_m = 4 \exp \left(\frac{V}{18}\right)\), \(\alpha_h = 0.07 \exp {\left(\frac{V}{20}\right)}\), \(\beta_h = \frac{1}{\exp\left(0.1(V+30)\right)+1}\).